


About Article
Investigation of inhibitory potential of resveratrol to the microtubule affinity regulating kinase 4: Implications in anticancer therapy
Abstract
Resveratrol shows efficacy against a wide range of cancers and neurodegeneration by targeting specific molecular pathways involved in cancer progression. It exhibits neuroprotective properties, which are crucial for reducing neurodegenerative diseases like Alzheimer's disease (AD). It works by reducing neuroinflammation, protecting neurons from oxidative damage, and controlling important signaling pathways that are essential for synaptic plasticity and neuronal survival. One of the main targets in cancer and neurodegenerative diseases, the kinase MARK4 is linked to the formation of tauopathies like AD. Resveratrol is an excellent antioxidant, anti-cancer and a neuroprotectant used as an inhibitor against the kinase MARK4. To comprehend the binding and structural changes in the protein's structure upon ligand binding, in silico investigations were conducted. After the in silico experiments, the pure protein was studied in vitro. With an IC50 value of 7 μ M, the natural substance resveratrol effectively inhibited the protein. Additionally, a significant binding affinity for MARK4 was demonstrated by the fluorescence binding experiment (K = 104 M−1). The findings demonstrated that the ligand attaches to the protein's binding pocket and blocks MARK4's enzymatic activity, indicating that the substance is a strong binding agent against MARK4.
Keywords
1. Introduction
Protein kinases form one of the largest and functionally most diverse enzyme groups of eukaryotic biology, which coordinate cellular signalling by reversible phosphorylation of serine, threonine, or tyrosine residues on substrate proteins (Mohammad et al., 2021; BIOVIA, 2017). It is a post-translational modification that functions as a molecular switch regulating protein activity, localisation, stability, and intermolecular interactions, and thus determining cellular fate choices (DeLano, 2002). Over 500 kinases constitute the human kinome, which mediates the combination of extracellular signals with intracellular events and controls cell proliferation, cell metabolism, cell cytoskeleton, apoptosis, and stress-adaptation (Atiya & others, 2023). Considering this pivotal regulatory position, disruptions in kinase network signaling have a close relationship with disease pathogenesis, especially cancer, in which disrupted phosphorylation cascades result in malignant transformation and resistance to therapy (Alrouji & others, 2023).
The development of cancer is defined by the constant activation of signaling pathways that maintain the proliferation, inhibit cell death, and induce invasion and metastasis (Dahiya & others, 2019). Although it has been widely studied, there is growing interest in kinases that control cellular architecture and polarity; many of these have been neglected due to their lack of association with the classical oncogenic kinases, including receptor tyrosine kinases and components of the mitogen-activated protein kinase MAPK pathway (Fan et al., 2019). These kinases not only provide structural support, but they are also involved in plasticity signalling, cell cycle and metastatic dissemination signalling. The microtubule affinity-regulating kinase (MARK) plays a role in cell cycle progression and cytoskeletal dynamics. (Shamsi & others, 2020).
MARK kinases are a subfamily of AMP-activated protein kinases (AMPK), and it comprises of four isoforms (MARK1-4) which regulate microtubule stability by phosphorylating microtubule-associated proteins (MAPs). MARK kinases regulate cytoskeletal reorganisation, vesicle trafficking, polarity, and organisation of mitotic spindles by regulating the separation of MAPs from microtubules (Anwar & others, 2022). MARK4 isoform that has received specific attention among them is its rampant tissue distribution and up-and-coming functions in the pathological conditions like those associated with neurodegenerative illnesses, metabolic imbalance, and malignancy. MARK4 is structurally a conserved catalytic kinase molecule with a regulatory region on either side of a conserved catalytic domain that enables it to participate in the coordination of structural and signaling pathways (Shamsi & others, 2020).
Research suggests that MARK4 is overexpressed in multiple types of cancers, and it leads to tumour progression in a variety of ways. Increased levels of MARK4 have been linked to increased cell proliferation, migration, invasion, and epithelial-to-mesenchymal transition (EMT)(Valeur & Berberan-Santos, 2013). Mechanistically, MARK4 controls cytoskeletal dynamics, controls mitotic spindle activity, and is linked with signalling cascades that regulate cellular survival and metabolic adjustment. This dualism structural organization MARK4 and signal transduction modulation makes this kinase a central non-canonical oncogenic kinase (Eftink, 1997; Lakowicz & Weber, 1973). Notably, the MARK4 activity seems to support metastatic phenotypes, such as epithelial polarity loss, motility, and stress-resistance, which can be regarded as its therapeutic importance.
Specific inhibition of kinases has significantly improved the therapeutic intervention of cancer, but the difficulties to overcome, including the development of resistance, off-target toxicity, and redundancy of pathways, remain. This means that the discovery of new targets of kinases and the development of other forms of inhibition methods are still priorities (Lakowicz, 2006). The reason why MARK4 is a promising candidate is that its blocking disrupts both cytoskeletal structure and oncogenic signalling pathways. Empirical evidence has shown that MARK4 blockade activates cell-cycle arrest, inhibits malignant cell proliferation, and induces apoptosis, which indicates that MARK4 blockers could provide multifunctional therapeutic effects (Vivian & Callis, 2001).
Along with the growth of the kinase-based drug discovery, natural products have resurfaced as a leading modulator of cellular signalling pathways (Taraska & Zagotta, 2010). Natural products offer structurally diversified chemical scaffolds that may interact with protein kinases in an ATP-competitive, substrate-competitive, or allosteric-mechanism [16]. The idea of natural molecules is also translational, with a number of anticancer drugs being based on the molecules that can be found in nature. Phytochemicals like flavonoids, phenolic acids, and terpenoids have been reported to control phosphorylation networks, dampen oncogenic signaling and increase cellular responses to stress in recent years [17]. Their pleiotropic property enables them to modulate many pathways at once, a property that is highly important in complex diseases such as cancer.
Natural compounds have been singled out by emerging research to have a specific MARK4 modulatory effect [18]. The research by Anwar et al. revealed that rosmarinic acid has anticancer effects due to its mechanism by which it directly interacts with MARK4 and consequently inhibits the proliferation of tumour cells and triggers apoptosis [19]. This study was a mechanistic understanding of the ability of phytochemical scaffolds to disrupt cytoskeletal kinase activity. Later studies indicated that the flavonoid myricetin suppresses breast and lung cancer cell growth by suppressing MARK4, which supports the use of the kinase as a therapeutic target [20]. Notably, these results indicated that structurally different natural molecules accumulate around MARK4, thereby giving the indication that the kinase has structural properties that can be utilized in the design of phytochemical-based drugs [21]. In addition to the sphere of oncology, MARK4 inhibition with the help of natural compounds has been shown in neurodegenerative conditions[22]. Bacopaside II was also discovered to mediate with MARK4 and regulate disease-related signalling, hence providing momentum on the broader biological importance of MARK4 targeting [18]. These studies, in general, define MARK4 as a key molecular hub involved in a variety of pathologies and, therefore, support the hypothesis that natural compounds could be universal modulators of this kinase. The targeting of cross-diseases in this way is especially appealing, as the control of cytoskeleton is central to cellular homeostasis [18].
Resveratrol (RV), a non-flavonoid polyphenol is a prevalent phytochemical under intense research in the field of bioactive plant-based compounds that demonstrates a comprehensive therapeutic capability. It is a derivative of stilbene that is found in grapes, berries, and peanuts having antioxidant, anti-inflammatory, cardioprotective, neuroprotective, and anticancer properties. [23]. On the molecular level, resveratrol disrupts a wide variety of signalling cascades, such as PI3K/Akt, MAPK, NF-κB signalling, p53, and sirtuin signalling. Such interactions regulate the major cell functions, including proliferation, apoptosis, angiogenesis and metabolic adaptation. All these interactions are shown to modulate cell proliferation, apoptosis, angiogenesis and metabolic adaptation processes which are the key processes in cancer biology [24].
RV has been demonstrated to cause tumor growth, cell cycle arrest, apoptosis and metastasis inhibition in cancer models. Its mechanism of action can often be interfered with by kinase-mediated phosphorylation cascades, and this implies that kinase modulation is a fundamental part of its anticancer effect [25]. It is also interesting to note that resveratrol can influence cytoskeletal organization and microtubule dynamics, which are closely linked to the MARK4 activity [22]. Resveratrol is a logical extension of the same mechanistic paradigm, which has been shown to be inhibited by related polyphenols, including myricetin and rosmarinic acid. It interacts with kinase catalytic domains via its polyphenolic structure and its pleiotropic signaling effects could increase its therapeutic activity by producing a coexisting effect on multiple pathways [26].
MARK4 is a relevant model of a kinase with its inhibition having the potential to modulate structural and signalling elements of tumour progression, and natural products, including RV, represent chemically versatile templates that can implement such changes [22]. The convergence highlights that phytochemical-derived kinase inhibitors will be useful as complements or adjuncts to current targeted therapies [30]. Recent research achievement has shown that it is possible to inhibit MARK4 and reduce the growth of cancer cells using natural compounds, which provide a mechanistic basis to phytochemical therapeutic interventions [31]. A prospective candidate in this emerging structure is resveratrol with its thoroughly established anticancer action and ability to regulate kinase signaling. The decoding of the interaction between MARK4 signalling and RV-mediated regulation could provide new opportunities for multi-target cancer therapy and precision medicine. The further convergence of kinase biology, structural pharmacology and natural product research is set to play a critical role in turning these findings into clinically relevant interventions.
2. Materials and Methods
2.1. Materials
Difco LB broth (Becton, Dickinson and Company, Sparks, MD, USA) was autoclaved at 121 ^°C for 15 min and stored at room temperature. IPTG (Sigma-Aldrich, Bengaluru, India; now Merck KGaA, Darmstadt, Germany) was prepared as a 1 M stock in Milli-Q water and stored at -20 ^°C. Kanamycin (50 mg/mL) and ampicillin (100 mg/mL) stock solutions were prepared in Milli-Q water and stored at -20 ^°C. CAPS buffer was prepared as a 1 M stock in Milli-Q water, and pH was adjusted as required. NaCl (5 M) and glycerol (100% v/v) stock solutions were prepared in Milli-Q water and stored at room temperature. Resveratrol (Sigma-Aldrich) was prepared as stock solutions in an appropriate solvent (e.g., DMSO) and stored according to the manufacturer’s instructions.
2.2. Molecular Docking
The three-dimensional crystal structure of human MARK4 was retrieved from the RCSB Protein Data Bank (PDB ID: 5ES1) at a resolution of 2.8 Å. Prior to docking, the protein structure was prepared by removing co-crystallised ligands and water molecules, followed by the addition of hydrogen atoms and assignment of Kollman charges using MGL Tools to ensure proper geometry and charge distribution for molecular docking analysis (Mohammad et al., 2021). Molecular docking of MARK4 with resveratrol was carried out using InstaDock. Blind docking was carried out to explore the surface of the entire protein, applying exhaustiveness of 8. The resultant poses were further analysed on the basis of binding affinity and pattern of interaction. All the interactions between the protein and ligand, including hydrogen bonds and hydrophobic interactions, were studied with Discovery Studio Visualizer and PyMOL (BIOVIA, 2017; DeLano, 2002).
2.3. Protein expression and purification
The protein MARK4 was expressed in M15 E.coli bacterial cells. The M15 strain contains the pREP4 plasmid, which has the lac repressor (lacI) gene, allowing regulation of protein expression under the T5 promoter in pQE vectors. The plasmid vector selected was pQE30 which is compatible with M15 and carried N-terminal polyHistidine tag, facilitating as a tag in purification and high yield. The recombinant plasmid with the protein of interest was transformed into E.coli-M15 cells, and protein purification was carried out with the previously established protocols (Atiya & others, 2023; Alrouji & others, 2023).
2.4. Enzyme inhibition assay
Enzyme here is a kinase, MARK4, which works by using ATP as a substrate for its activity, converting ATP to ADP, and the released phosphate is used by the kinase to phosphorylate its target. For the assay mentioned here, the principle is that the protein kinase is considered 100 per cent active by measuring the amount of phosphates that are released upon interaction with ATP. Keeping the concentration of protein constant, different concentrations of ligand are added to the protein and incubated, allowing the ligand to bind with the protein. Afterwards, ATP is added to the control, i.e. protein without ligand and protein with increasing concentration of ligand, and the released phosphate is estimated with a reagent known as BIOMOL green reagent. MARK4 and ATP stock solutions were prepared, and the reaction was set in a 96-well plate by mixing protein, ATP, buffer and ligands to obtain a final MARK4 concentration of 5 μ M and ATP of 200 μ M. The reaction mixtures were first set up without ATP and incubated with increasing concentrations of ligands for 30 minutes at 37^°C. Further, ATP was added and incubated at 37^°C for approximately 30 minutes. Following incubation, Biomol Green reagent was added according to the manufacturer’s instructions to terminate the reaction and enable detection of released inorganic phosphate. The mixtures were thoroughly mixed and incubated at room temperature in the dark to allow full color development. Absorbance was measured at 620 nm using a microplate reader.
2.5. Fluorescence binding studies
The binding studies of RV with MARK4 were performed on the Jasco spectrofluorometer at 25^°C. The compound was prepared by dissolving it in DMSO, giving the desired 1 mM working concentration in the working buffer (pH 8.0). The binding studies were performed with a fixed concentration of MARK4 (4 μ M), and the ligand was added gradually in increasing concentration till saturation. The addition of ligand was mixed thoroughly and gently to ensure interaction between protein and ligand before measurements. The intrinsic fluorescence of protein, mainly contributed by the aromatic amino acids and predominantly Tryptophan, was measured by excitation at 280 nm, which generated an emission spectrum at 300–400 nm. The fluorescence quenching observed in the emission spectra was analysed using the modified Stern-Volmer equation (Dahiya & others, 2019).
In the given equation, F0 and F denote the fluorescence intensity of protein and protein ligand complex, respectively. K is the binding constant; n denotes number of binding sites and C for concentration of RV.
fracF0F = 1 + Ksv[C]
logleft(fracF0 - FFright) = log K + n log [C]
2.6. Cell proliferation assay
Cancer cells H1299 are a widely used human non-small cell lung carcinoma (NSCLC) cell line, specifically identified as a large cell carcinoma subtype. The cells were procured from NCCS, Pune, India. A culture medium was prepared for H1299 cells with antibiotics and heat-inactivated FBS. Using a 96-well culture plate, approximately 5 × 103 cells were plated per well following passages till passage number 20, with a confluency of 85%, and all the treatments were given. A compound stock solution was made with DMSO, with a maximum limit of 0.2% per treatment. The MTT stock was prepared as 5 mg/ml, and 10 μ L of the stock was incubated in all the treated wells at 37^°C with 5% CO2 for a few hours. The 96-well plate was read spectrophotometrically at 570 nm using a 96-well plate reader.
3. Results and Discussion
3.1. Molecular docking analysis
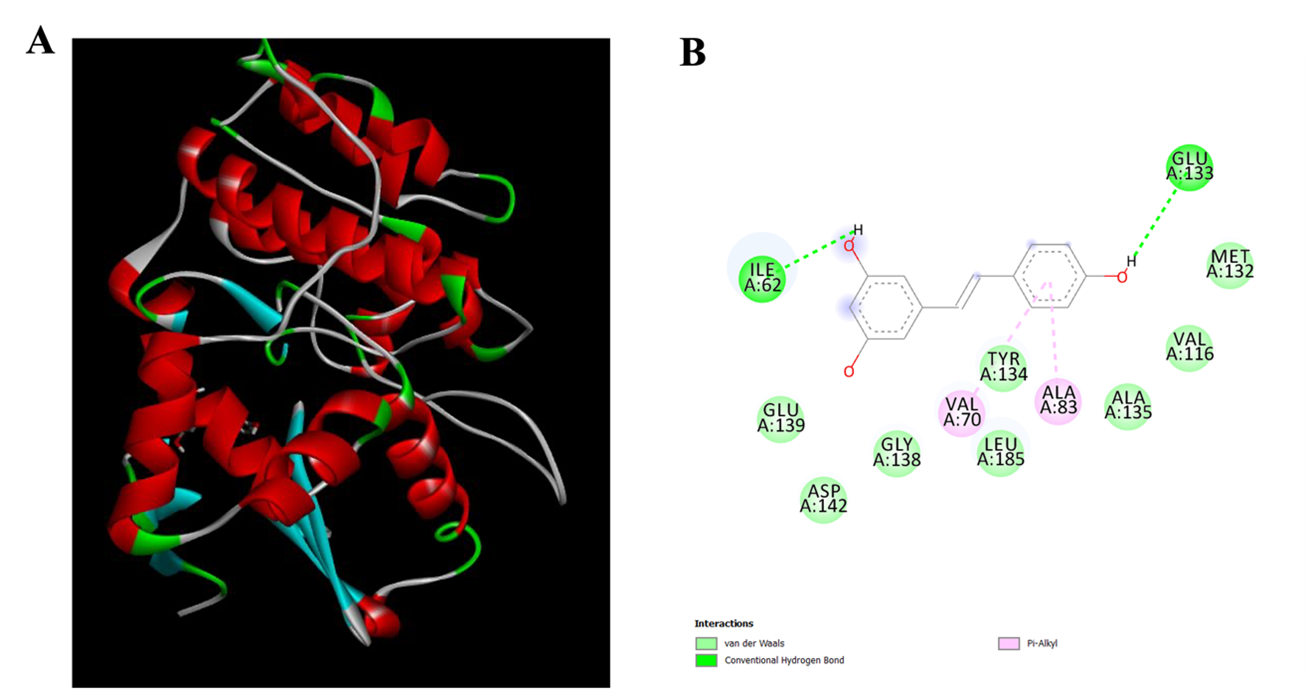
To understand the binding of RV with MARK4, molecular docking studies were carried out as shown in Figure 1. According to the docking research, RV interacts with Ile62 and Glu133 by typical hydrogen bonds, which are crucial for anchoring the ligand inside the binding cavity. The stability of ligand-protein complexes is greatly aided by hydrogen bonds, which are important factors in determining ligand specificity. RV also forms multiple hydrophobic and van der Waals interactions with several residues lining the MARK4 binding pocket, including Met132, Val116, Tyr134, Gly138, Glu139, Asp142, Ala135, and Leu185. Together, these interactions help keep the ligand inside the kinase's catalytic domain and stabilize the structure of the MARK4–RV complex. Moreover, π–alkyl interactions with Val70 and Ala83 were noted, suggesting stronger hydrophobic interactions between RV's aromatic ring and the nonpolar residues in the active site. Docking studies reveal RV occupies the catalytic pocket of MARK4 with multiple interactions (Fan et al., 2019; Shamsi & others, 2020; Anwar & others, 2022).
3.2. Fluorescence-based binding studies
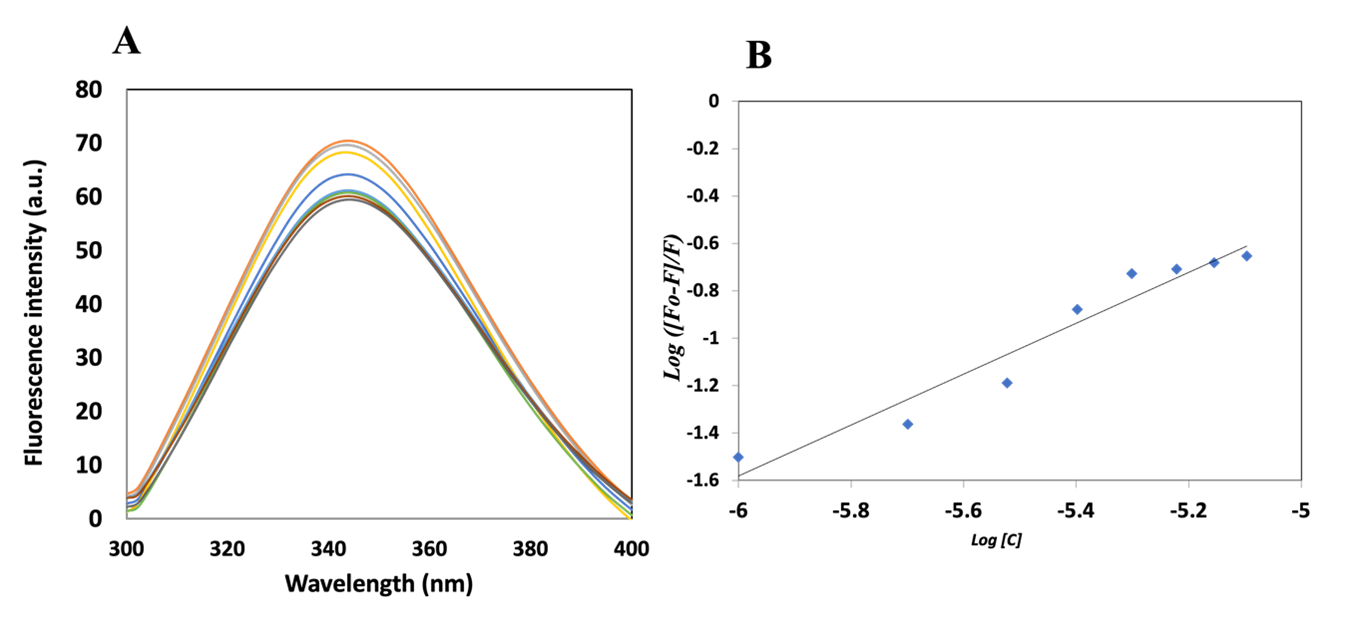
The aromatic amino acids in the protein structure are responsible for the intrinsic fluorescence of the protein. The major contributor amongst all aromatic amino acids is tryptophan (Trp), followed by tyrosine and phenylalanine. Trp has a high quantum yield, resulting in high protein fluorescence at 300–350 nm when excited at 280 nm. The intensity and spectral position of this emitted light provide information about the structural and environmental status of the protein (Valeur & Berberan-Santos, 2013; Eftink, 1997; Lakowicz & Weber, 1973). Trp fluorescence intensity and emission wavelength can be dramatically changed by changes in solvent polarity, conformational shifts, or ligand binding, which makes it an effective probe for researching protein structure, folding, and intermolecular interactions (Lakowicz, 2006; Vivian & Callis, 2001). As tryptophan fluorescence is highly sensitive to conformational rearrangements and binding-induced perturbations, it is widely used to investigate protein-ligand interactions, conformational changes, and to estimate binding constants (Taraska & Zagotta, 2010). In the current investigation, an increase in the ligand concentration resulted in decreased fluorescence intensity of the protein, as shown in Figure 2A. The quenching in the fluorescence of the protein can likely be due to the changes in the microenvironment near the tryptophan residues in the presence of RV. The binding constant of the interaction was estimated to be 7.0 × 104 M−1 using the modified Stern-Volmer equation (Figure 2B).
3.3. Enzyme inhibition assay
The kinase MARK4 is an important therapeutic target due to its association with various diseases, including cancer and neurodegeneration. MARK4 has a strong correlation with Alzheimer’s disease, as MARK4 is associated with pathological phosphorylation of tau protein, contributing to dementia and early onset of AD. MARK4 is upregulated in various cancers, including breast cancer, lung cancer and hepatocellular carcinoma (Mohammad et al., 2021; BIOVIA, 2017; DeLano, 2002). Therefore, MARK4 has emerged as an interesting therapeutic target in cancer, neurodegeneration and other diseases. RV is a naturally occurring compound with documented anti-inflammatory, anti-oxidant, anti-ageing, anti-cancer and neuroprotective activities. In order to study the influence of RV on kinase activity and to complement molecular docking and fluorescence-based binding studies, an enzyme inhibition assay was carried out.
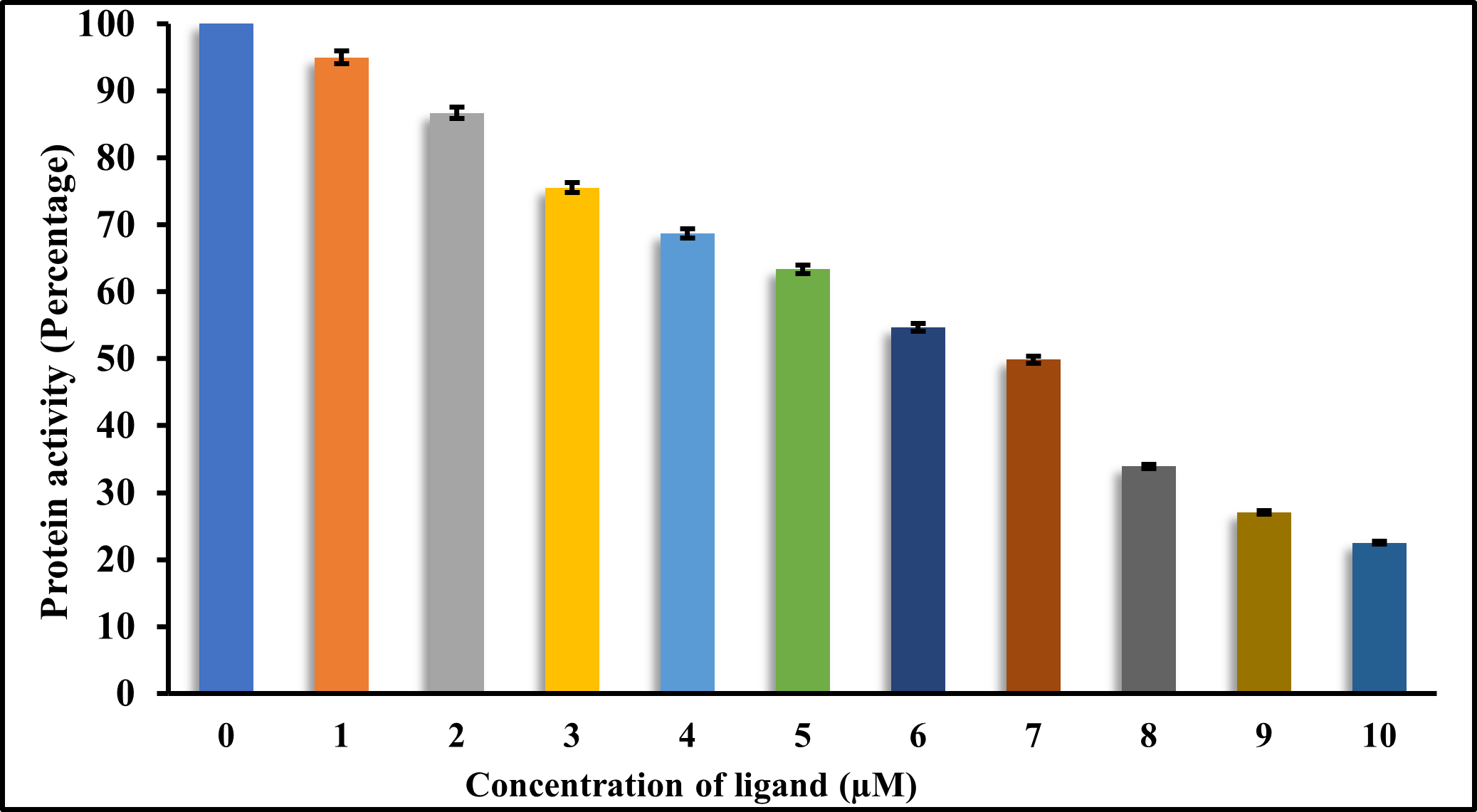
A decrease in kinase activity of MARK4 was observed with increasing ligand concentration (Figure 4), indicating inhibition of the kinase activity of the protein, which can be due to the formation of protein-ligand association, as evident from docking studies. AAT Bioquest IC50 calculator was used to determine the half-maximal inhibitory concentration (IC50), which is the inhibitor concentration needed to decrease enzyme activity by 50% as shown in Figure 3. In line with earlier findings, the IC50 value for RV was found to be approx 7 μ M. (Dahiya & others, 2019; Fan et al., 2019). These findings validate RV's therapeutic potential and confirm its status as a strong MARK4 inhibitor. RV might be a promising addition to the expanding family of small-molecule kinase inhibitors that target MARK4, considering the pivotal role that kinases play in a variety of pathological situations, such as cancer and neurological diseases.
3.4. Cell proliferation assay
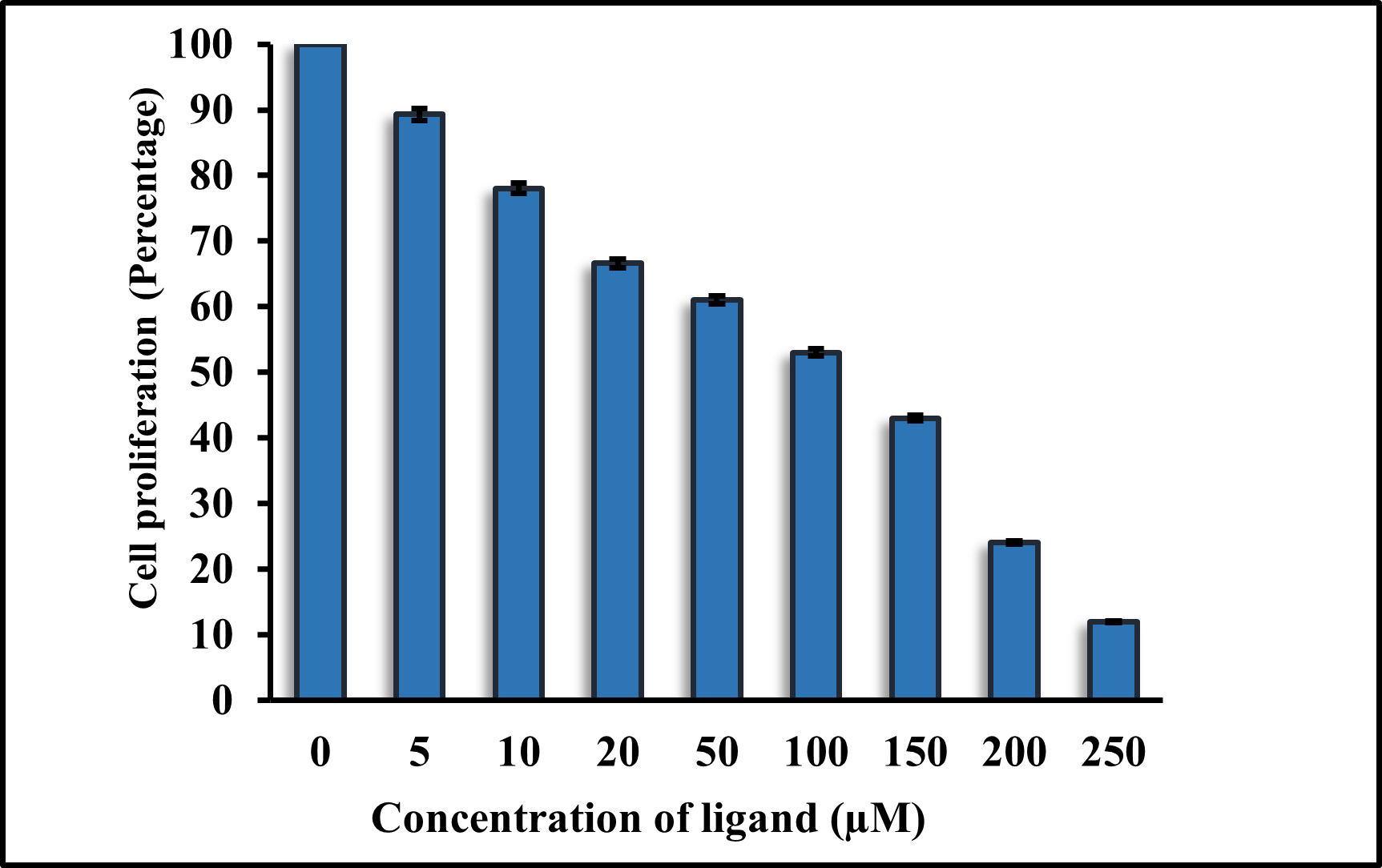
H1299 lung cancer cell lines were used to examine the cytotoxicity of RV against them. We performed a cell survival assay to verify the anti-cancerous activity of the compounds. The cell growth was observed for 48 hours following the treatment with increasing concentrations of RV. Compared to the DMSO vehicle control, RV showed significant cytotoxicity in the H1299 cell lines. The IC50 values of RV against H1299 were approx 114 μ M, as shown in Figure 4.
4. Conclusion
To conclude, the current paper presents an in-depth analysis of resveratrol as an antimicrotubular affinity-regulating kinase 4 (MARK4) inhibitors, a kinase that is gradually gaining prominence as the cause of cancer advancement and neurodegenerative diseases. Combining both computational and experimental methods, we can show that resveratrol forms a strong interaction with MARK4 and that it can effectively regulate the enzyme activity. The molecular docking analysis exhibited positive binding of resveratrol in the catalytic site of MARK4, which is an indication of structural compatibility and may dislodge the kinase activity. The binding studies conducted using fluorescence also supported the presence of a direct interaction between resveratrol and MARK4 as demonstrated by the quenching of intrinsic protein fluorescence with rising resveratrol concentration, and calculation of a strong binding constant, which indicates a high affinity. Enzyme inhibition assays of functional validation showed that resveratrol inhibits the activity of the MARK4 kinase dose-dependently with an IC50 of the order of micromolar, which is an indicator of a good small-molecule inhibitor.
In combination, these data indicate resveratrol as a promising phytochemical scaffold that could be used to target MARK4 and disrupt the kinase-mediated signalling pathways related to cytoskeletal regulation, tumour progression and neurodegeneration. The additional multi-target signalling capabilities of resveratrol enhance the future of resveratrol as a systems-level therapeutic agent. Nevertheless, more research in cellular models, structural dynamics and pharmacokinetic optimization is required to convert these results into clinical applications. Although these encouraging results exist, there are still translation issues. Naturally occurring substances tend to have low bioavailability, short metabolism, and situational potency. These limitations are being overcome using medicinal chemistry, structural biology, and delivery systems based on nanotechnology to optimise the scaffold of phytochemicals and enhance pharmacokinetic activities. In-depth knowledge of the interactions of MARK4 with ligands, such as binding site structure and conformational dynamics, will be fundamental to rational drug design [29].
Altogether, this article contributes to the comprehension of MARK4 modulation by natural compounds and offers a mechanistic concept of phytochemical-based kinase inhibitors evolution. The MARK4 resveratrol-interaction is also a good beginning in rational drug design to multi-pathway therapeutic intervention of cancer and other related diseases.
Declaration of Competing Interest
The authors report no conflicts of interest.
Data availability statement
All data generated or analyzed during this study are included in this article.
Figures
Figure 1. Molecular docking of RV with MARK4; A. 3D interaction of RV and MARK4; B. 2D interactions of RV and MARK4.
{kind=link}
Figure 2. (A) Fluorescence emission spectra of native MARK4 protein and increasing concentration of RV (0–8 μ M). (B) Modified Stern-Volmer plot of MARK4–RV system.
{kind=link}
Figure 3. ATPase activity assay of MARK4 with varying RV concentration (0–10 μ M).
{kind=link}
Figure 4. Cell proliferation assay of RV against cell line H1299.
{kind=link}
References
- 1
Mohammad, T., Mathur, Y., Hassan, M. I. 2021, Briefings in Bioinformatics, 22
- 2
BIOVIA, Dassault Systemes 2017, Discovery Studio Visualizer
- 3
DeLano, W. L. 2002, CCP4 Newsletter on Protein Crystallography, 40, 82–92
- 4
Atiya, A., & others 2023, Journal of Biomolecular Structure and Dynamics, 41, 8824–8830
- 5
Alrouji, M., & others 2023, International Journal of Biological Macromolecules, 235, 123831
- 6
Dahiya, R., & others 2019, RSC Advances, 9, 23302–23315
- 7
Fan, J., Fu, A., Zhang, L. 2019, Quantitative Biology, 7, 83–89
- 8
Shamsi, A., & others 2020, Biomolecules, 10, 789
- 9
Anwar, S., & others 2022, Journal of Molecular Liquids, 355, 118928
- 10
Valeur, B., & Berberan-Santos, M. N. 2013, Molecular Fluorescence: Principles and Applications, (John Wiley & Sons)
- 11
Eftink, M. R. 1997, Methods in Enzymology, 278, 221–257
- 12
Lakowicz, J. R., & Weber, G. 1973, Biochemistry, 12, 4161–4170
- 13
Lakowicz, J. R. 2006, Principles of Fluorescence Spectroscopy, (Springer)
- 14
Vivian, J. T., & Callis, P. R. 2001, Biophysical Journal, 80, 2093–2109
- 15
Taraska, J. W., & Zagotta, W. N. 2010, Neuron, 66, 170–189